

At Which Of The Following Stages Does Transcriptional Control Occur?
If our genes are then similar, what really makes a eukaryote dissimilar from a prokaryote, or a human from E. coli? The reply lies in the divergence in gene expression and regulation used.
It is estimated that the human genome encodes approximately 25,000 genes, about the same number as that for corn and virtually twice every bit many equally that for the common fruit fly. Even more interesting is the fact that those 25,000 genes are encoded in about 1.v% of the genome. So, what exactly does the other 98.5% of our Deoxyribonucleic acid do? While many mysteries remain virtually what all of that extra sequence is for, nosotros know that information technology does contain complex instructions that direct the intricate turning on and off of gene transcription.
Eukaryotes Require Complex Controls Over Gene Expression
While basic similarities in factor transcription exist between prokaryotes and eukaryotes—including the fact that RNA polymerase binds upstream of the gene on its promoter to initiate the process of transcription—multicellular eukaryotes command prison cell differentiation through more complex and precise temporal and spatial regulation of gene expression.
Multicellular eukaryotes take a much larger genome than prokaryotes, which is organized into multiple chromosomes with greater sequence complexity. Many eukaryotic species carry genes with the aforementioned sequences every bit other plants and animals. In improver, the aforementioned DNA sequences (though not the aforementioned proteins) are constitute inside all of an organism's diploid, nucleated cells, even though these cells form tissues with drastically different appearances, properties, and functions. Why then, is there such great variation among and inside such organisms? Quite merely, the way in which different genes are turned on and off in specific cells generates the diverseness we find in nature. In other words, specific functions of different cell types are generated through differential gene regulation.
Of grade, higher eukaryotes still respond to environmental signals by regulating their genes. Merely there is an additional layer of regulation that results from cell-to-cell interactions within the organism that orchestrate development. Specifically, factor expression is controlled on ii levels. Beginning, transcription is controlled by limiting the amount of mRNA that is produced from a detail gene. The 2d level of command is through post-transcriptional events that regulate the translation of mRNA into proteins. Fifty-fifty after a poly peptide is made, mail-translational modifications can touch its activity.
Transcriptional Regulation in Eukaryotes
Regulation of transcription in eukaryotes is a result of the combined effects of structural properties (how Deoxyribonucleic acid is "packaged") and the interactions of proteins called transcription factors. The virtually important structural departure between eukaryotic and prokaryotic DNA is the formation of chromatin in eukaryotes. Chromatin results in the different transcriptional "ground states" of prokaryotes and eukaryotes (Tabular array i).
Tabular array 1: Overview of Differences Betwixt Prokaryotic and Eukaryotic Gene Expression and Regulation
| Prokaryotes | Eukaryotes | |
| Structure of genome | Unmarried, generally circular genome sometimes accompanied by smaller pieces of accompaniment DNA, like plasmids | Genome found in chromosomes; nucleosome construction limits Deoxyribonucleic acid accessibility |
| Size of genome | Relatively small | Relatively large |
| Location of gene transcription and translation | Coupled; no nucleoid envelope barrier because of prokaryotic cell structure | Nuclear transcription and cytoplasmic translation |
| Gene clustering | Operons where genes with similar part are grouped together | Operons generally non found in eukaryotes; each gene has its ain promoter element and enhancer element(south) |
| Default country of transcription | On | Off |
| DNA structure | Highly supercoiled DNA with some associated proteins | Highly supercoiled chromatin associated with histones in nucleosomes |
Transcription Factors and Combinatorial Control
Transcription factors (TFs) are regulatory proteins whose function is to activate (or more rarely, to inhibit) transcription of Dna by binding to specific DNA sequences. TFs have defined Deoxyribonucleic acid-binding domains with upward to 106-fold higher affinity for their target sequences than for the residue of the DNA strand. These highly conserved sequences have been used to categorize the known TFs into various "families," such as the MADS box-containing proteins, SOX proteins, and POU factors (Remenyi et al., 2004). Transcription factors can also be classified by their three-dimensional poly peptide construction, including basic helix-turn-helix, helix-loop-helix, and zinc finger proteins. These unlike structural motifs result in transcription gene specificity for the consensus sequences to which they bind.
Sequence-specific transcription factors are considered the nearly important and diverse mechanisms of gene regulation in both prokaryotic and eukaryotic cells (Pulverer, 2005). In eukaryotes, regulation of gene expression by transcription factors is said to exist combinatorial, in that it requires the coordinated interactions of multiple proteins (in contrast to prokaryotes, in which a single protein is usually all that is required).
Many genes, known as housekeeping genes, are needed by about every type of prison cell and appear to be unregulated or constitutive. But at the cadre of cellular differentiation, manifested in the diversity of prison cell types observed in different organisms, is the regulation of gene expression in a tissue-specific manner. The aforementioned genome is responsible for making the entire cadre of cell types, each of which has its own function—for example, red blood cells exchange oxygen, muscle cells aggrandize and contract, and cells in the immune organization recognize pathogens. Genes that regulate cell identity are turned on under very specific temporal, spatial, and ecology weather to ensure that a cell is able to perform its designated function.
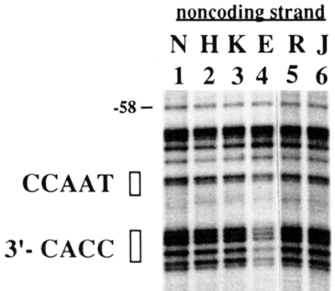
Accept the example of the cistron for beta globin, a protein used in reddish blood cells for oxygen exchange. Every cell in the human body contains the beta globin factor and the corresponding upstream regulatory sequences that regulate expression, just no cell type other than red blood cells expresses beta globin. Scientists can utilise a technique called DNA footprinting to map where transcription factors bind to specific regulatory sequences. When Reddy et al. examined the beta globin promoter in different cell types, they establish that the transcription factors that could demark to the promoter sequences required for beta globin expression were expressed but in erythroblasts (immature adult red claret cells). (See Effigy 1). The 2 consensus sequences in the beta globin promoter known for binding transcription factors, CCAAT and CACC, were protected in the erythroid cells (E), merely non the other jail cell types (Reddy et al., 1994).
The "Ground State" of Deoxyribonucleic acid Expression
RNA polymerase in prokaryotes can admission almost any promoter in a Deoxyribonucleic acid strand without the presence of activators or repressors. Thus, the "ground country" of Deoxyribonucleic acid expression in prokaryotes is said to be nonrestrictive, or "on." In eukaryotes, however, the ground state of expression is restrictive in that, although strong promoters might be present, they are inactive in the absence of some sort of recruitment to the promoter by transcription factors. For instance, RNA polymerase 2, which transcribes mRNA, cannot bind to promoters in eukaryotic Dna without the aid of transcription factors (Struhl, 1999). In many eukaryotic organisms, the promoter contains a conserved gene sequence called the TATA box. Various other consensus sequences also exist and are recognized past the dissimilar TF families. Transcription is initiated when one TF binds to one of these promoter sequences, initiating a series of interactions betwixt multiple proteins (activators, regulators, and repressors) at the same site, or other promoter, regulator, and enhancer sequences. Ultimately, a transcription complex is formed at the promoter that facilitates binding and transcription by RNA polymerase.
As in prokaryotes, eukaryotic repressor molecules tin can sometimes demark to silencer elements in the vicinity of a gene and inhibit the bounden, associates, or activity of the transcription complex, thus turning off expression of a gene. Positive regulation by TFs that are activators is common in eukaryotes. Considering the restrictive transcriptional ground country, it is logical that positive regulation is the predominant form of command in all systems characterized to engagement. Many activating TFs are mostly bound to DNA until removed by a betoken molecule, while others might just bind to Deoxyribonucleic acid once influenced past a signal molecule. The bounden of ane type of TF can influence the bounden of others, as well. Thus, gene expression in eukaryotes is highly variable, depending on the type of activators involved and what signals are present to command binding.
The Role of Chromatin
Even when transcription factors are present in a cell, transcription does not always occur, because often the TFs cannot reach their target sequences. The association of the Dna molecule with proteins is the get-go pace in its silencing. The associated Dna and histone proteins are collectively called chromatin; the complex is tightly bonded by allure of the negatively charged DNA to the positively charged histones (Tabular array ane). The state of chromatin can limit access of transcription factors and RNA polymerase to Deoxyribonucleic acid promoters, contributing to the restrictive ground land of gene expression. In social club for cistron transcription to occur, the chromatin structure must exist unwound.
Chromatin structure contributes to the varying levels of complexity in gene regulation. It allows simultaneous regulation of functionally or structurally related genes that tend to be nowadays in widely spaced clusters or domains on eukaryotic Dna (Sproul et al., 2005). Interactions of chromatin with activators and repressors can issue in domains of chromatin that are open up, closed, or poised for activation. Chromatin domains have various sizes and different extents of stability. These variations allow for phenomena found solely in eukaryotes, such as transcription at diverse stages of evolution and epigenetic memory throughout cell division cycles. They also let for the maintenance of differentiated cellular states, which is crucial to the survival of multicellular organisms (Struhl, 1999).
Multiple Interactions Provide Synchronous Control
Every bit you have seen, the state of chromatin structure at a specific region in eukaryotic DNA, forth with the presence of specific transcription factors, works to regulate factor expression in eukaryotes. However, this complex coaction betwixt proteins that serve as transcriptional activators or repressors and accessibility to the regulatory sequence is notwithstanding just part of the story. Epigenetic mechanisms, including DNA methylation and imprinting, noncoding RNA, mail service-translational modifications, and other mechanisms, further enrich the cellular portfolio of factor expression command activities.
References and Recommended Reading
Pulverer, B. Sequence-specific DNA-binding transcription factors. Nature Milestones (2005) doi: 10.1038/nrm1800 (link to commodity)
Reddy, P. M., Stamatoyannopoulos, G., Papayannopoulou, T., & Shen, C. Chiliad. Genomic footprinting and sequencing of human beta-globin locus: Tissue specificity and cell line artifact. Journal of Biological Chemical science 269, 8287–8295 (1994)
Remenyi, A., Scholer, H., & Wilmanns, M. Combinatorial control of gene expression. Nature Structural and Molecular Biology xi, 812–815 (2004) doi:10.1038/nsmb820 (link to article)
Struhl, Chiliad. Fundamentally dissimilar logic of gene regulation in eukaryotes and prokaryotes. Cell 98, 1–4 (1999)
Sproul, D., Gilbert, N., & Bickmore, W. The part of chromatin structure in regulating the expression of clustered genes. Nature Reviews Genetics 6, 775–781 (2005) doi:10.1038/nrg1688 (link to article)

At Which Of The Following Stages Does Transcriptional Control Occur?,
Source: https://www.nature.com/scitable/topicpage/regulation-of-transcription-and-gene-expression-in-1086/
Posted by: framptontunt1941.blogspot.com
0 Response to "At Which Of The Following Stages Does Transcriptional Control Occur?"
Post a Comment